










Россия
Культивирование судака в личиночный период развития является одним из наиболее сложных этапов в технологии. Неоднородность роста личинок и их высокая смертность препятствовали возможности разработать или использовать существующие модели для описания роста судака на ранних этапах развития. Решение проблемы стартового питания судака с использованием искусственных кормов позволило получить относительно стабильные результаты выращивания с возможностью моделирования роста судака в период раннего постэмбрионального развития. Целью работы является оценка адекватности существующих моделей, способных описать рост личинок судака в условиях индустриальной аквакультуры, а также разработка и проверка собственной модели. Было исследовано 8 нелинейных моделей, которые применяются в аквакультуре для математического описания ростовых процессов – трех- и четырехпараметрические логистические модели, модели фон Берталанфи, Шнуте, Гомпертца, Михаэлиса – Ментена и Вейбулла, а также собственная экспериментальная линейная модель. Основой для проверки адекватности моделей были фактические данные о росте личинок судака, полученные в ходе их выращивания в бассейнах полностью на искусственных кормах. Эффективность и адекватность моделей определяли при помощи информационного критерия Акаике AIC, байесовского информационного критерия BIC, оценки среднеквадратичной ошибки MSE и стандартной ошибки, а также времени итерации. В результате исследований было показано, что рост судака на личиночном этапе развития лучшим образом моделируют четырехпараметрические модели – логистическая и экспериментальная линейная модель, а также модель фон Берталанфи с тремя параметрами. При этом четырехпараметрические модели имели лучшие показатели AIC, BIC и коэффициента R2, тогда как время итерации было лучшим у трехпараметрической модели. Остальные модели показали значительно худшие результаты оценки и на фоне нереалистичных прогнозов роста не могут быть использованы для описания роста личинок судака. Полученные результаты позволят лучше понять закономерности роста ранних личинок судака, помогут скорректировать технологию его выращивания и повысить эффективность разведения.
аквакультура, судак, личинки, математическая модель роста рыбы, итерация, количество параметров
Введение
Показатели роста рыбы являются наиболее значимыми аспектами экономической эффективности рыбоводства. Крайне важно получать данные о росте в режиме реального времени с целью корректировки параметров выращивания, таких, например, как нормы кормления. В настоящее время, чтобы получить данные о длине и весе гидробионтов, соответствующие измерения в аквакультуре по-прежнему производятся вручную, что легко может вызвать стресс и травмы у рыб, особенно на ранних стадиях развития.В аквакультуре применяются функции роста, отражающие рост рыб на определенной стадии культивирования, такие как абсолютный рост, относительный рост и удельная скорость роста. Данные функции базируются на эмпирических данных и не позволяют прогнозировать дальнейший рост рыб [1]. Однако на их основе возможно построение уравнений, обобщающих информацию из эмпирических данных о росте в набор биологически интерпретируемых параметров [2]. Модели роста с такими параметрами были использованы для многих видов рыб: тюрбо Scophthalmus maximus [1], нильской тиляпии Oreochromis niloticus [2], африканского сома Clarias gariepinus [3], групера Epinephelus moara [4], радужной форели Oncorhynchus mykiss [5] и др. Но следует учитывать существующие различия в темпах роста рыб на разных стадиях их онтогенеза, что требует использования нескольких моделей для описания роста одного вида рыб в течение жизни. Например, для радужной форели описано 3 стадии роста, характерные для мелких (менее 20 г), средних (20–500 г) и крупных рыб (более 500 г), существенно отличающихся друг от друга [6]. В целом рост рыб можно разделить на две фазы: ювенильную фазу, в которой вся избыточная энергия используется для соматического роста, и фазу после полового созревания, в которой часть или вся избыточная энергия направляется на воспроизводительную функцию [7]. Lester et al. [8] разработали модель роста, основанную на этих двух фазах: рост неполовозрелых рыб был описан с помощью линейной модели, в то время как рост половозрелых особей следует модели роста фон Берталанфи. До сих пор большинство исследований моделей в основном базировались на данных о росте «товарной» рыбы, т. е. от молоди до взрослых особей, в то время как для ранней молоди рыб, особенно с очень мелкими личинками, такими как у судака, использование моделей роста сильно ограничено. Судак востребован во многих сферах рыбного хозяйства – для восполнения численности природных популяций [9], поддержки коммерческого и любительского рыболовства [10], использования в качестве биомелиоратора [11] и др. Продолжает расти интерес к коммерческому выращиванию судака [12]. Высокая востребованность судака в рыбохозяйственной отрасли делает весьма актуальной разработку модели роста этого вида, однако информация о темпах роста судака ограниченна [13]. Одним из самых сложных этапов в технологии культивирования судака является выращивание его личинок от начала питания внешней пищей. Несмотря на обширную информацию об описании роста личинок судака, представленную в литературе, создание модели роста на основе этих данных ранее не представлялось возможным. Это связано с неоднородностью роста судака на ранних личиночных этапах, сопровождающейся высокой смертностью, в том числе от каннибализма, и отсутствием рабочей технологии выращивания личинок. В комплексе это не позволяло создать модель роста судака в ранний период жизни. Полученные в последние годы положительные результаты в области разработки специализированных стартовых искусственных кормов для судака [14], а также стабильные результаты его выращивания от начала питания личинок до сеголеток (собственные материалы) позволяют провести анализ существующих и разработать собственную модель для описания и прогнозирования роста судака в аквакультуре, что и явилось целью данной работы.
Материал и методы исследования
Сбор материала о росте личинок судака проводили на рыбоводном хозяйстве ООО «Форват» (оз. Суходольское, Приозерский р-н., Ленинградская обл.) в 2022–2024 гг. Икру получали от маточного стада, выращенного в садках на искусственных кормах. После инкубации и становления «на плав» (трое суток после вылупления) личинок рассаживали в квадратные производственные бассейны площадью 4 м2 при плотности посадки 50 экз./л. От начала питания и до достижения массы около 150 мг личинок кормили только искусственным кормом Rusander (ООО «Русло», Россия). Воду в бассейны подавали из природного водоисточника – оз. Суходольского. Личинок выращивали в естественном температурном режиме. В период исследований вода плавно прогревалась от 17 до 21 °C. Размерно-весовые параметры личинок определяли во время контрольных обловов, проводимых на 6, 9, 13, 21, 38 и 48 сутки после вылупления. В разные годы рост личинок был примерно одинаков, в связи с чем для работы с моделями использовали усредненные данные показателей роста. Для сравнения и систематического анализа существующих биологических моделей роста были выбраны 8 моделей, которые применяются в аквакультуре для математического описания ростовых процессов – трех- и четырехпараметрические логистические модели (М1 и М2 соответственно), модели Гомпертца (М4), фон Берталанфи (М5), Вейбулла (М6), Михаэлиса – Ментена (М9) и Шнуте (М10) (все модели были взяты из [4, 15, 16]), а также собственная экспериментальная полулинейная динамическая модель с обратной связью (М11). Уравнения расчета и количество используемых в моделях параметров представлены в табл. 1, где G представляет собой максимальное расчетное значение модели; G(t) – рассчитываемая величина прироста за время; G0 – начальный вес рыбы; t – возраст рыбы, сут; а и b – параметры скорости, которые определяют, насколько быстро достигается G; с и g – параметры формы; D – асимптотический вес во времени t; в модели Шнуте t1 и t2 представляют собой начальный и конечный возраст соответственно, а y1 и y2 – расчетный вес в возрасте t1 и t2 соответственно; as – постоянное значение относительной скорости роста; Inc – среднесуточный прирост.
Таблица 1
Table 1
Модели роста для прогнозирования прироста судака на ранних стадиях развития
Growth models for predicting the growth of walleye in the early stages of development
|
Модель |
Уравнение |
Количество |
|
M1 – логистическая |
Gt = G / [1 + a · exp(–b · t)] |
3 |
|
М2 – логистическая |
Gt = G + (G0 – G) / (1+(t / D)c) |
4 |
|
М4 – Гомпертца |
Gt = G · exp(–a · exp(–b · t)) |
3 |
|
M5 – фон Берталанфи |
Gt = G + (G0 – G) (1 – exp (–b · t))c |
3 |
|
М6 – Вейбулла |
Gt = (G – G0) exp(–(b · t)c) |
4 |
|
М9 – Михаэлиса – Ментена |
Gt = (G0 · Dc + G · tс) / (Dc + tc) |
4 |
|
M10 – Шнуте |
Gt = y1 + (y2 – y1) ((1 – exp(–as (t – t1))) / |
3 |
|
M11 – линейная модель |
G = Gt + Gt+1 · Inc 0,01 |
4 |
При выборе модели предпочтение отдается ее успешному применению в схожих контекстах и, что наиболее важно, предполагаемой точности прогнозирования. Для этого на основе расчетов моделей были построены графики роста личинок, которые сравнивали с фактическим ростом судака, также выраженным в виде графика. Построение графиков и подгонку моделей осуществляли в Python (3.6.13), который является широко используемым программным обеспечением для моделирования с множеством функций и методов подгонки.
С целью выбора оптимальной модели, адекватно описывающей рост личинок судака, были применены следующие критерии оценки: среднеквадратичная ошибка (MSE), информационный критерий Акаике (AIC) и байесовский информационный критерий (BIC), коэффициент R2, время итерации. Данные параметры были рассчитаны с помощью пакета «scipy.stats» в Python. Уравнения данных параметров представлены в соответствии с [4, 5]:
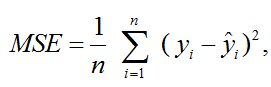
где n – объем данных; yi – исходные данные; ŷi – прогнозируемые данные;
где k – количество параметров в модели; L – максимальное значение функции правдоподобия, n – размер выборки.
Для оценки надежности моделей было подсчитано время итераций. В моделировании итерация – это повторение определенной последовательности действий (например, этапов разработки или вычислений) для постепенного улучшения или достижения более точного результата. Итеративный подход в моделировании позволяет решать сложные системы пошагово, добавляя или совершенствуя функции с каждым циклом, что делает процесс более гибким и эффективным, снижая риски и повышая качество конечной модели. Таким образом, время итерации – это промежуток времени, в течение которого модель реализует задачу.
Модель с наименьшими расчетными значениями исследованных показателей является наиболее подходящей для прогнозирования и моделирования роста личинок судака в индустриальной аквакультуре.
Также был рассчитан коэффициент R2, который показывает долю дисперсии зависимой переменной относительно независимых переменных в модели и вычисляется уравнением
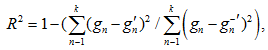
где  – наблюдаемые значения;
– наблюдаемые значения; ![]() – значения, предпредсказанные моделью;
– значения, предпредсказанные моделью;  – среднее из наблюдаемых значений;
– среднее из наблюдаемых значений; 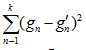 – сумма квадратов остатков (разница между наблюдаемыми и предсказанными значениями);
– сумма квадратов остатков (разница между наблюдаемыми и предсказанными значениями); 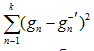 – общая сумма квадратов (разница между наблюдаемыми значениями и их средним); R2 находится в диапазоне от 0 до 1 (1 означает, что модель идеально объясняет дисперсию, 0 – что модель не объясняет дисперсию зависимой переменной).
– общая сумма квадратов (разница между наблюдаемыми значениями и их средним); R2 находится в диапазоне от 0 до 1 (1 означает, что модель идеально объясняет дисперсию, 0 – что модель не объясняет дисперсию зависимой переменной).
Результаты
Результатом исследования моделей стало построение графиков роста личинок судака (рис.), которое выполнено на основе данных о массе тела личинок, полученных расчетным путем: по оси ординат – масса, мг, по оси абсцисс – сутки эксперимента.
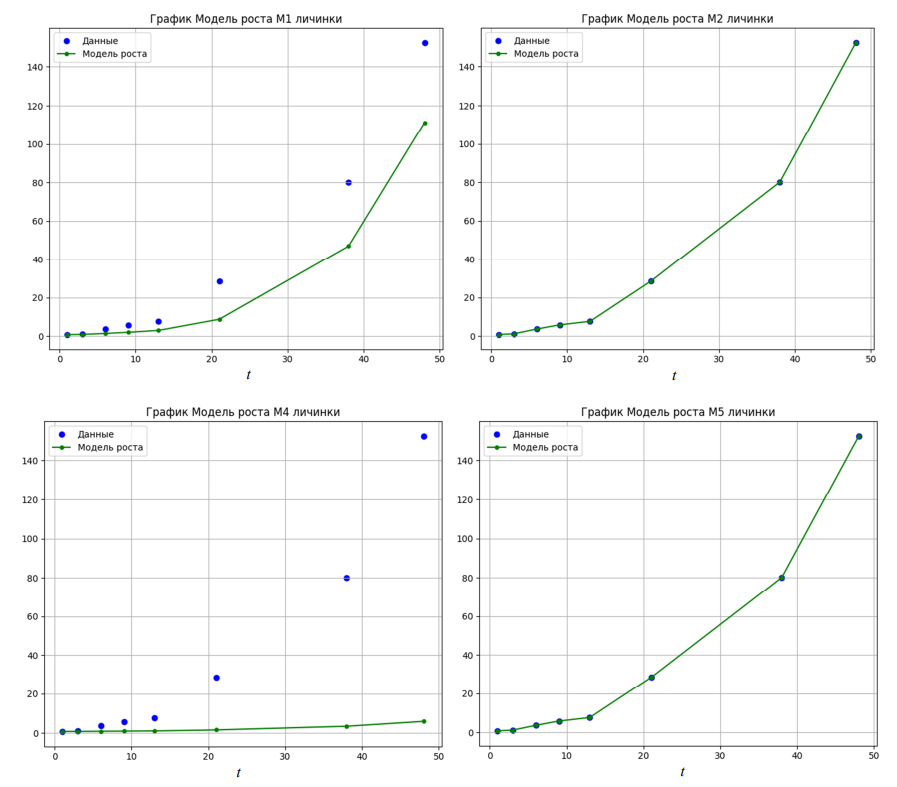
Кривые весового роста личинок судака, построенные в соответствии с моделями роста
Weight growth curves of walleye larvae, constructed in accordance with the growth models
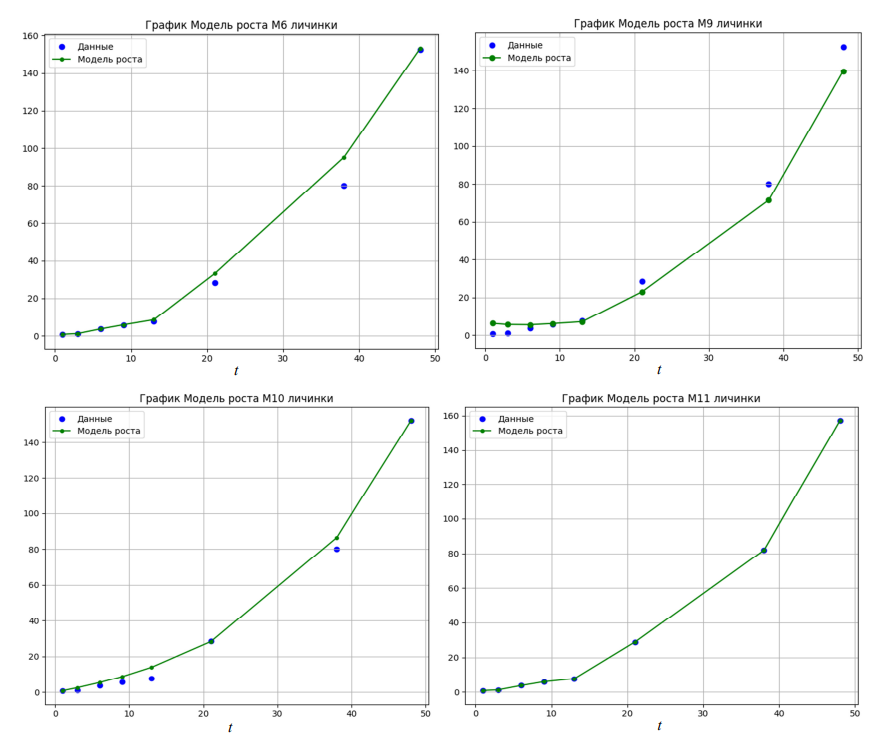
Окончание рис. Кривые весового роста личинок судака, построенные в соответствии с моделями роста
Ending Fig. Weight growth curves of walleye larvae, constructed in accordance with the growth models
Наиболее точно рост личинок судака описывают несколько моделей – четырехпараметрическая логистическая модель (М2), модель фон Берталанфи (М5) и собственная модель роста (М11) (см. рис.). Трехпараметрическая модель Гомпертца (М4) не способна адекватно описать динамику роста судака на личиночной стадии развития: личинки к концу периода выращивания достигают массы около 8 мг, что в 19 раз ниже фактической конечной массы личинок, равной 152 мг. Модель Вейбулла (М6) незначительно завышает промежуточные показатели роста личинок с 13 по 40-е сутки, но адекватно рассчитывает конечную индивидуальную массу судака, близкую к фактической. Модель Михаэлиса – Ментена (М9) в первые 13 суток выращивания демонстрирует отсутствие прироста у личинок, затем описывает рост судака кривой, имеющей вид, схожий с фактическим графиком роста, но с небольшим отставанием. Кривая роста, построенная в соответствии с трехпараметрической моделью Шнуте (М10), в конечной точке пересекается с фактическими данными, однако в промежуточных значениях часто показывает завышенный прирост веса. Собственная экспериментальная полулинейная динамическая модель с обратной связью (М11), использующая для расчетов 4 параметра, при построении кривой весового роста личинок судака показала полное соответствие фактическим данным (см. рис.). Оценка моделей по выбранным критериям показывает, что наименьшие значения AIC, BIC и MSE соответствуют четырехпараметрической логистической модели (М2). Также у этой модели максимальное значение R2, но относительно высокое время итерации: в 2 раза выше, чем у модели фон Берталанфи (М5). В свою очередь, модель М5 характеризуется высокими показателями AIC, BIC и MSE
и низким R2 (табл. 2).
Таблица 2
Table 2
Критерии оценки для моделей весового роста личинок судака
Evaluation criteria for weight growth models of walleye larvae
|
Модель |
Критерии |
||||
|
AIC |
MSE |
R2 |
Итерация, с |
||
|
М1 |
69,85 |
70,01 |
5,84 |
0,45 |
0,0110 |
|
М2 |
–321,16 |
–320,93 |
0 |
1 |
0,0206 |
|
М4 |
–276,61 |
–276,45 |
2,14 · 10–8 |
1 |
0,0602 |
|
М5 |
69,85 |
70,01 |
61,31 |
0,31 |
0,0104 |
|
М6 |
56,79 |
56,95 |
27,10 |
0,71 |
0,0294 |
|
М9 |
–302,84 |
–302,69 |
4,68 · 10–9 |
1 |
0,0143 |
|
М10 |
21,57 |
21,97 |
4,2488 |
0,99 |
0,0021 |
|
М11 |
43,67 |
43,83 |
142,48 |
0,96 |
0,0253 |
Остальные модели показали значительно худшие результаты оценки и на фоне нереалистичных прогнозов роста не могут быть использованы для описания роста личинок судака.
Обсуждение
Быстрый рост и укороченная продолжительность личиночной фазы позволяют ранней молоди рыб быстрее выйти из-под пресса хищников, что обеспечивает успешное выживание и восполнение природных популяций [17]. Известно, что рост – это функция, которая находится в зависимости от условий среды обитания организма. Важными факторами, влияющими на рост и развитие личинок, являются температура и доступность пищи [18]. У теплолюбивых рыб, к которым относится судак, обитание в теплой воде стимулирует более высокую скорость метаболизма [19], что приводит к ускорению роста [20]. В условиях аквакультуры оба стимулирующих рост фактора могут находиться на уровне оптимума. Это обеспечивается быстрым прогревом воды до благоприятных для роста личинок температур, что происходит в первую неделю после вылупления (с учетом периода, когда личинки/предличинки не питаются), а также избыточным кормлением, характерным для ранней молоди рыб. Известно, что искусственные корма менее эффективны в реализации потенциала роста личинок, чем живые корма. Но темп роста личинок судака на искусственном рационе значительно выше, чем у старших возрастных групп. Например, среднесуточный прирост трехлеток судака в благоприятных температурных условиях (20–22 °С) на искусственном корме не превышает 1,2 %, что практически на порядок ниже (а в некоторые периоды и больше), чем у личинок, получавших сухой корм. Для описания столь быстрого и неоднородного роста мы использовали различные нелинейные модели. Нелинейные модели могут выражать биологические характеристики роста на основе задаваемых параметров. Эти параметры, имеющие биологическую значимость, в результате работы модели формируют кривую роста. Модели, выбранные в данном исследовании, характеризуются как трех- и четырехпараметрические, значения параметров которых описывают биологический рост. Например, G определяет максимальный предел роста рыбы, G0 – начальный рост, b – скорость роста, а c влияет на форму кривой роста и делает модель более гибкой. Наиболее реалистично личиночный рост судака описывали две модели с четырьмя параметрами (М2 и М11) и одна трехпараметрическая модель (М5). Недавние исследования на групере показали, что рост личинок этого вида может быть успешно смоделирован с помощью моделей с четырьмя параметрами, в то время как все трехпараметрические модели не смогли дать достоверных оценок прироста, а в некоторых случаях дальнейшие характеристики кривой роста были нереалистичными [4]. Следует отметить, что исследование роста групера в ранний период развития является единственной работой в доступной литературе, в которой исследуется рост ранних личинок в аквакультуре. Полученные нами данные отчасти согласуются с результатами моделирования роста групера, за исключением того, что в нашем случае трехпараметрическая модель фон Берталанфи показала адекватный прогноз роста личинок судака.
В целом при исследовании роста личинок лучшим образом себя реализуют модели с бóльшим количеством параметров (если сравнивать трех- и четырехпараметрические модели). Это подтверждается другими исследованиями роста рыб в аквакультуре. Так, Baer et al. [21] использовали четырехпараметрическую модель Шнуте для подгонки роста тюрбо Psetta maxima (от 5 до 2 000 г) и обнаружили, что это оптимальная модель с реалистичным коэффициентом роста. Lugert et al. [22] также подтвердили, что четырехпараметрическая модель имеет преимущества при подгонке роста рыб и обладает большим потенциалом для моделирования роста в аквакультуре. Однако, с одной стороны, предполагается, что увеличение числа параметров приведет к более высокому качеству соответствия, с другой стороны, может иметь противоположный эффект, увеличивая время итерации, т. е. привести к «переподгонке» [23]. Вероятно, этим можно объяснить увеличение времени итерации для эффективных моделей с четырьмя параметрами (М2 и М11) по сравнению с трехпараметрической моделью (М5). Таким образом, на основе проведенных исследований можно заключить, что рост судака на личиночном этапе развития лучшим образом моделируют четырехпараметрические модели – логистическая (М2) и экспериментальная полулинейная динамическая модель с обратной связью (М11), а также модель фон Берталанфи с тремя параметрами (М5). При этом четырехпараметрические модели имели лучшие показатели информационного критерия Акаике (AIC), байесовского информационного критерия (BIC) и коэффициента R2, тогда как время итерации было лучшим у трехпараметрической модели.
Заключение
В последнее время моделирование приобретает все более значимое влияние на управление аквакультурой. Особенно это важно для сектора рыбоводства, связанного с выращиванием личинок и молоди рыб. В данной работе исследован рост судака в аквакультуре в первый год выращивания. Показано, что из 8 нелинейных моделей роста, в том числе широко используемых в аквакультуре, для прогнозирования роста личинок судака подходят 3 модели – четырехпараметрические логистическая и экспериментальная полулинейная динамическая модель с обратной связью и трехпараметрическая модель фон Берталанфи. При этом лучшим критериям статистической оценки соответствуют модели с четырьмя параметрами, что в комплексе с возможностью адекватно прогнозировать рост личинок делает их наиболее перспективными в аквакультуре судака. Однако другой ключевой параметр для оценки моделей – время итерации – был минимальным у трехпараметрической модели, что может быть связано с меньшим количеством параметров в модели и, как следствие, более быстрым достижением конечного результата. Необходимо подчеркнуть, что работа над исследованием роста судака и моделированием его процессов с использованием математических функций стала возможной благодаря решению проблемы стабильного получения жизнестойкого физиологически полноценного материала в виде личинок и сеголеток судака. Полученные результаты позволят лучше понять закономерности роста ранней молоди судака и помогут скорректировать технологию его выращивания и повысить эффективность разведения.
1. Lugert V., Thaller G., Tetens J., Schulz C., Krieter J. A review on fish growth calculation: multiple functions in fish production and their specific application // Reviews in Aquaculture. 2016. V. 8 (1). P. 30–42. https://doi.org/10.1111/raq.12071.
2. Zardin A. M. S. O., de Oliveira C. A. L., de Oliveira S. N., Yoshida G. M., de Albuquerque D. T., de Campos C. M., Ribeiro R. P. Growth curves by Gompertz nonlinear regression model for male and female Nile tilapias from different genetic groups // Aquaculture. 2019. V. 511. P. 734243. https://doi.org/10.1016/j. aquaculture.2019.734243.
3. Musa B. O., Hernández-Flores A., Adeogun O. A., Oresegun A. Determination of a predictive growth model for cultivated African catfish Clarias gariepinus (Burchell, 1882) // Aquaculture Research. 2021. V. 52 (9). P. 4434–4444. https://doi.org/10.1111/are.15282.
4. Sun X., Wang Y. Growth models in aquaculture for hybrid and natural groupers based on early development stage // Aquaculture. 2024. V. 578 (3). P. 740026. https://doi.org/10.1016/j.aquaculture.2023.740026.
5. Коновальчикова E. Н., Ивашко Е. Е. Подбор параметров моделей роста радужной форели // Вестн. Астрахан. гос. техн. ун-та. Сер.: Рыбное хозяйство. 2024. № 2. С. 42–48. https://doi.org/10.24143/2073-5529-2024-2-42-48.
6. Dumas A., France J., Bureau D. P. Evidence of three growth stanzas in rainbow trout (Oncorhynchus mykiss) across life stages and adaptation of the thermal-unit growth coefficient // Aquaculture. 2007. V. 267 (1–4). P. 139–146. https://doi.org/10.1016/j. aquaculture.2007.01.041.
7. Day T., Taylor P. D. Von Bertalanffy’s growth equation should not be used to model age and size at maturity // The American Naturalist. 1997. V. 149. P. 381–393.
8. Lester N. P., Shuter B. J., Abrams P. A. Interpreting the von Bertalanffy model of somatic growth in fishes: the cost of reproduction // Proceedings of the Royal Society B: Biological Sciences. 2004. V. 271. P. 1625–1631. DOIhttps://doi.org/10.1098/rspb.2004.2778.
9. Mustamäki N., Bergström U., Ådjers K., Sevastik A., Mattila J. Pikeperch (Sander lucioperca (L.)) in decline: high mortality of three populations in the northern Baltic Sea // Ambio. 2014. V. 43 (3). P. 325–336.
10. Bninska M., Wolos A. Management of selected polish commercial and recreational lake fisheries activities // Fisheries Management and Ecology. 2001. V. 8 (4–5). P. 333–343.
11. Королёв А. Е. Оценка биомелиоративной роли двухлеток судака (на примере оз. Гусиного) // Сб. науч. тр. ГосНИИОРХ. 1989. Т. 292. С. 51–59.
12. Lappalainen J., Dorner H., Wysujack K. Reproduction biology of pikeperch (Sander lucioperca (L.)) – a review // Ecology of freshwater fish. 2003. V. 12 (2). P. 95–106.
13. Dalsgaard J., Lund I., Thorarinsdottir R., Drengstig A., Arvonen K., Pedersen P. B. Farming different species in RAS in Nordic countries: current status and future perspectives // Aquacultural Engineering. 2013. V. 53. P. 2–13. https://doi.org/10.1016/j.aquaeng.2012.11.008.
14. Лютиков А. А., Барышников А. В., Королев А. Е., Трифонов А. Е., Шумская Н. В., Новиков В. Ю. Применение продуктов микробиосинтеза и их гидролизатов в качестве источника доступного белка в стартовых кормах для судака // Актуальные проблемы освоения водных биологических ресурсов Российской Федерации: материалы Всерос. конф. ученых и специалистов, посвящ. 160-летию Н. М. Книповича (Мурманск, 27–28 октября 2022 г.). Мурманск: Изд-во ПИНРО им. Н. М. Книповича, 2023. С. 331–337.
15. Panik M. J. Fundamentals of Population Dynamics // Growth Curve Modeling: Theory and Applications. John Wiley & Sons, 2014. P. 352–371.
16. Gottschalk P. G., Dunn J. R. The five-parameter logistic: a characterization and comparison with the four-parameter logistic // Analytical biochemistry. 2005. V. 343. P. 54–65. https://doi.org/10.1016/j.ab.2005.04.035.
17. Houde E. D. Recruitment variability: in Fish reproductive biology / eds. T. Jakobsen, M. J. Fogarty, B. A. Megrey, E. Moksness. Oxford, UK: Blackwell Publishing Ltd., 2009. P. 91–171.
18. Pörtner H. O., Peck M. A. Climate change effects on fishes and fisheries: Towards a cause-and-effect understanding // J. Fish. Biol. 2010. V. 77. P. 1745–1779. DOIhttps://doi.org/10.1111/j.1095-8649.2010.02783.x.
19. Peck M. A., Reglero P., Takahashi M., Catalán I. A. Life cycle ecophysiology of small pelagic fish and climate-driven changes in populations // Prog. Oceanogr. 2013. V. 116. P. 220–245. DOIhttps://doi.org/10.1016/j.pocean.2013.05.012.
20. Houde E. D. Comparative growth, mortality, and energetics of marine fish larvae: temperature and implied latitudinal effects // Fish. Bull. 1989. V. 87. P. 471–495.
21. Baer A., Schulz C., Traulsen I., Krieter J. Analysing the growth of turbot (Psetta maxima) in a commercial recirculation system with the use of three different growth models // Aquaculture International. 2011. V. 19 (3). P. 497–511. https://doi.org/10.1007/s10499-010-9365-0.
22. Lugert V., Tetens J., Thaller G., Schulz C., Krieter J. Finding suitable growth models for turbot (Scophthalmus maximus L.) in aquaculture 1 (length application) // Aquac. Res. 2017. V. 48 (1). P. 24–36. https://doi.org/10.1111/are.12857.
23. Burnham K., Anderson D. Model selection and multimodel inference: a practical information-theoretic approach. London: Springer, 2002. 488 p.
